
按分类浏览
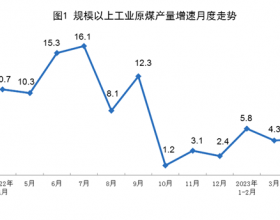

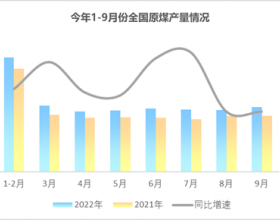
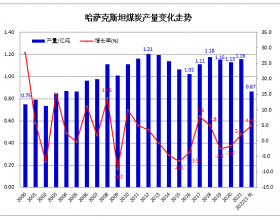
推荐新闻
最新价格行情
- [钢材价格] 2024-12-26全国主要城市H型钢价格汇总
- [钢材价格] 2024-12-26全国主要城市工字钢价格汇总
- [钢材价格] 2024-12-26全国主要城市槽钢价格汇总
- [钢材价格] 2024-12-26全国主要城市角钢价格汇总
- [钢材价格] 2024-12-26全国主要城市中厚板价格汇总
- [钢材价格] 2024-12-26全国主要城市冷轧板卷价格汇总
- [钢材价格] 2024-12-26全国主要城市热轧板卷价格汇总
- [钢材价格] 2024-12-26全国主要城市线材价格汇总
- [钢材价格] 2024-12-26全国主要城市螺纹钢价格汇总
- [钢材价格] 2024-12-25全国主要城市H型钢价格汇总